
The Australian Agronomist Magazine Summer 2019 issue features an article by Neutrog’s Microbiologist, Dr. Uwe Stroeher called ‘Carbon Dioxide and Water Use Efficiency.’ Uwe talks about the current economic and climatic conditions which are presenting some never before seen challenges to Australian agriculture, and how carbon dioxide and its availability plays a critical role in water use efficiency in plants.
Without a doubt, the current economic and climatic conditions are presenting some never before seen challenges to Australian agriculture. The most pressing issue is the availability of water, and how to efficiently use this limited resource.
Perhaps surprisingly, carbon dioxide (CO2) and its availability plays a critical role in water use efficiency in plants, and to understand this we need to look at how CO2 is obtained and used by various plants.
As a rule, plants take up CO2 via the stomatal pores in the leaves. These pores open and close according to various environmental signals, including the requirement for CO2. Interestingly, not all plants use CO2 for photosynthesis in the same way – in essence, commercially important plants can be divided in C3 and C4 plants depending on how they use CO2.
In C3 and C4 plants, the ‘C’ designates the carbon, whereas in CAM plants the ‘C’ stands for Crassulacean. These acronyms define the three ways in which plants fix CO2 during photosynthesis. Fixing carbon is the way plants remove the CO2 from atmosphere and turn it into organic molecules like carbohydrates (Figure 1).
C3 plants are plants in which the initial product of the assimilation of CO2 through photosynthesis is 3-phosphoglycerate, which contains 3 carbon atoms, whereas a C4 plant is a plant that fixes CO2 initially into a four-carbon sugar. C4 plants are thought to have evolved more recently, and are very efficient in hot, dry climates.
The C3 group of plants (many of which have evolved in cool, wet climates) include cereal crops such as wheat, barley and rye, and important commercial crops such as rice and cotton, as well as many vegetable and fruit varieties. In these plants, the use of CO2 occurs directly in the mesophyll cells of the leaves (mesophyll cells being the ones most responsible for photosynthesis), and the CO2 that is taken up by the leaves is used directly in the Calvin cycle (Figure 1).
Although this may sound efficient, it is only effective when the CO2 concentration is high within the cell. When the CO2 concentration drops below 50ppm, the plant may inadvertently uses oxygen instead of CO2 in a process known as photo-respiration. This is extremely wasteful and can lead to a drop in yield of up to 30%. Therefore, the presence of oxygen becomes a major inhibitor for efficient photosynthesis in C3 plants. In order to overcome this inefficiency, C3 plants keep their stomata open in order to take in more CO2, but this comes at a cost, as opening the stomatal pores allows water to escape, therefore leading to a drop in the water use efficiency of these plants¹.
As mentioned above, in C4 plants, the CO2 is fixed in the mesophyll cells to form a four-carbon sugar (C4) in a pathway known as the Hatch–Slack pathway (which is not affected by oxygen). This C4 substrate is shuttled to the bundle cells within the plant where it is converted to C3, and the CO2 is liberated and is ready to be used in the Calvin cycle away from any extraneous oxygen (Figure 1). In this manner, these plants don’t need to open their stomata to the same degree, as excess oxygen and lower CO2 is not an issue, so as a result, these plants lose less water and have a higher water use efficiency.
Although there are a number of commercially important C4 plants such as corn, sorghum, sugar cane and millet, the vast majority of plants are C3 plants, and as such there is no ready alternative to these in agriculture. There is a third group of plants called the CAM plants (Crassulacean Acid Metabolism) which include pineapples and succulents these plants are the most efficient users of water, as they only open their stomatal pores during the night to take in CO2. Similar to the C4 plants, these have a four-carbon intermediate which they produce at night. This is converted to a three-carbon substrate with the liberation of CO2 during the day, and the CO2 is then available for the Calvin cycle in a fashion similar to what occurs in the C4 plants (Figure 1).

An increase in the water use efficiency can benefit both C3 and C4 plants, so although C3 plants are the most affected by low CO2, C4 plants are not immune to water loss due to the stomatal pore opening triggered by low CO2 concentrations. In essence, the one way to overcome the issue of poor water use efficiency in plants is to increase the available CO2 (Figure 2 – source: https://phys.org/news/2017-07-carbon-dioxide-world-water-wise.html).
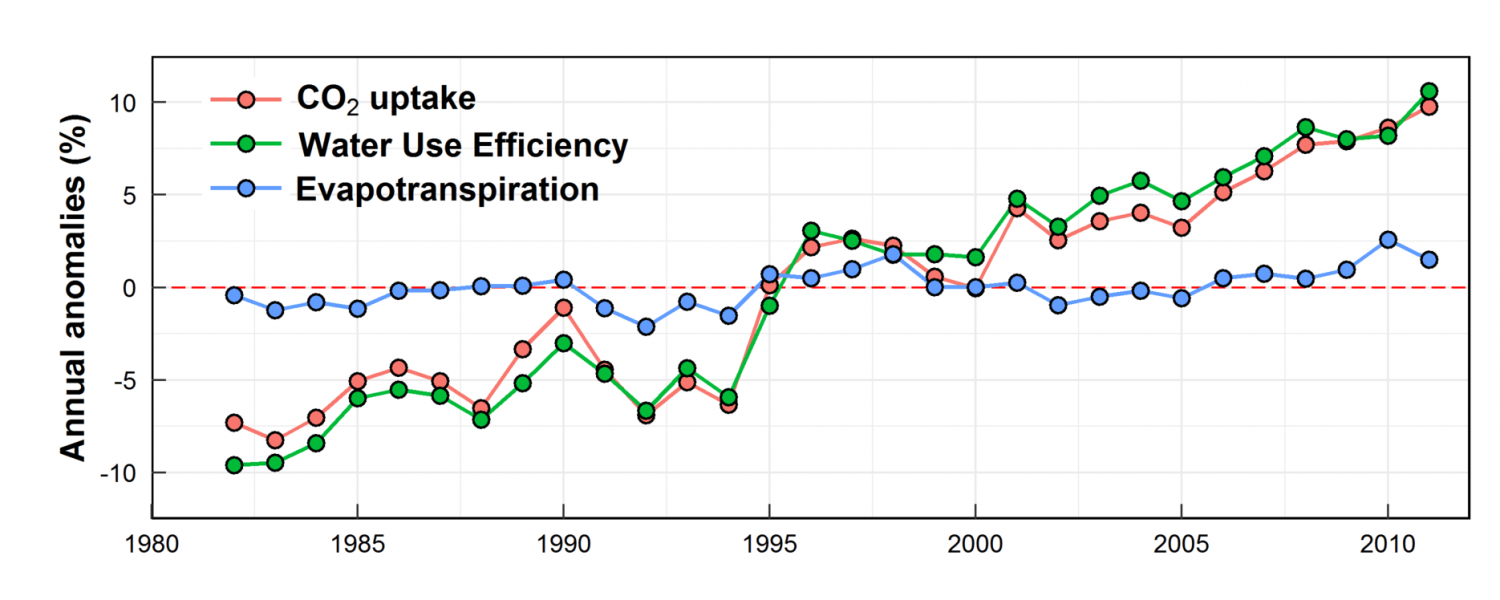
Higher levels of CO2 have been shown to significantly increase the growth of C3 plants and improve their water use – this is known as the ‘CO2 fertilisation effect’, where the result on C4 plants is less obvious when it comes to plant growth and yield. Some increases in water use efficiency have come about by the increase of CO2 in our atmosphere, which currently stands at about 400 ppm.³ This increase in water use efficiency may be in part due to partial closure of the stomata as well as increased levels of CO2, which results in fewer stomatal pores in the leaf. However, this increase in the greenhouse gas comes with its own problems, such as changing rainfall patterns, increases in average temperatures and drought, which negates much of the improvement in growth due to increasing CO2 levels. This appears to be especially obvious in Australia.³
In essence, the ideal situation would be to see an increase of the CO2 concentration in the vicinity of the plant, but not globally. The easiest and most cost-effective way to do this is to increase soil respiration rates by the use of organic inputs. Organic material (either plant or animal waste) consists of a significant proportion of organic carbon, which is carbon that is tied up in organic molecules, in substrates such as cellulose, lignin or oils, or more easily decomposed material such as starches, sugars and proteins.
When prokaryotic organisms such as bacteria and fungi metabolise these materials, they release CO2 which is immediately available for plants to use (Figure 3). In effect, what then occurs is a localised but not a global increase in CO2 concentration. The use of mulches or composts are ideal for this – we know that mulches will most obviously save water by reducing evaporative loses. We want to also increase the soil respiration rate, however using mulch with a high carbon to nitrogen ratio such as straw, woodchips and sugarcane tend to be biologically less active, and as such, will release much less CO2 compared to a mulch which has a balance of carbon and nitrogen. A mulch such as ‘Whoflungdung’ from Neutrog (which is manufactured by screening out the bedding material (straw and shavings) from composted broiler/poultry manure) is much more biologically active, and will therefore release significantly more CO2 than an inert mulch.
Composts incorporated into the soil will have a similar effect – during the breakdown of this material, CO2 will stream out of the ground to be used by plants. As CO2 is heavier than normal air, the plants which will benefit the most are those that are either low-growing such as vegetables and cereal crops, and obviously younger smaller plants.

Journal References:
1. CO2 sensing and CO2 regulation of stomatal conductance: advances and open questions. C. Engineer, M. Hashimoto-Sugimoto, J. Negi, M. Israelsson-Nordstrom T. Azoulay-Shemer,W-J. Rappel, K. Iba and J. Schroeder. Trends Plant Sci. 2016 (1):16-30. 2. Benefits of CO2 enrichment on crop plants are modified by soil water status. S. Kang, F. Zhang, X. Hu and J. Zhang. Plant and Soil. 2002 (238): 69-77. 3. Recent increases in terrestrial carbon uptake at little cost to the water cycle. L. Cheng, L. Zhang, Y-P. Wang, J.G. Canadell, F.H.S. Chiew, J. Beringer, L. Li, D.G. Miralles, S. Piao and Y. Zhang. Nature Communications 2017 (8) Article #: 110